Introduction
Neurosteroids are a type of steroid produced in the central nervous system rather than the adrenal gland. Neurosteroids rapidly alter neuronal excitability through interaction with neurotransmittergated ion channels of GABAA receptors.1,2 In addition, neurosteroids may also exert effects on gene expression via intracellular steroid hormone receptors. Neurosteroids have a wide range of potential clinical applications from sedation to epilepsy and traumatic brain injury treatment.3
Systemic administration of pilocarpine-induced status epilepticus (SE) consequentially generates death of pyramidal cells in the CA1 and CA3 fields of the hippocampus.4 This is due to excessive elevation of intracellular Ca2+ levels, resulting from overactivation of glutamate receptors.5,6 Therefore, regulation of intracellular Ca2+ levels is an important factor in neuronal survival after SE.
Calcium binding proteins, such as calbindin-D28k (CB), parvalbumin, and calretinin, are present in hippocampal non-pyramidal cells.7 It was reported that they have the capacity to buffer and transport intracellular Ca2+ in the nervous system.8,9 CB is ubiquitously expressed in the hippocampus, and particularly abundant in the granule cell-mossy fiber system.9,10 A number of studies have reported that CB has a neuroprotective function modulating intracellular Ca2+ levels against various central nervous system insults, including epilepsy.11,12
In previous studies, decreased CB expression was observed in epileptic conditions13, and some steroids, such as testosterone and estrogen, stimulated CB synthesis.14,15 However, the effect of neurosteroids on CB expression in epilepsy is unclear.
Therefore, we performed this study to examine whether neurosteroids, particularly androsterone, affect CB expression. We aimed to understand the relationship between the neuroprotective effects of androsterone and changes in CB expression with pilocarpine-induced SE.
Methods
The Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) approved all procedures. Male Institute for Cancer Research (ICR) mice (25 to 30 g, Orient Bio Co., Gyeonggi-do, Korea) were used in these experiments. Mice were housed under a 12 h light/dark cycle with food and water ad libitum. Lithium chloride (127 mg/kg, Sigma, St. Louis, MO, USA) was injected intraperitoneally (i.p) 24 h prior to pilocarpine administration. Next, mice were pretreated with methscopolamine-bromide (1 mg/kg, i.p., Sigma) 30 min prior to pilocarpine, and single dose of pilocarpine (30 mg/kg, i.p., Sigma) was administered. Seizures were scored using Racine’s scale, and the duration of status epilepticus (SE) was determined by behavioral assessment. The beginning of SE was defined as the onset of continuous generalized seizure activity (stage 4 or 5 based on Racine’s scale) without regaining normal behavior between the seizures. All mice received i.p. injections of 0.9% saline: 5 mL twice on the day with SE and 5 ml twice (in the morning and evening) of the day after SE. Two hours after SE, mice were treated with androsterone (100 and 200 mg/kg, i.p.). We compared with three other groups: the saline-only injection group (normal), the androsterone 200 mg-only injection group (androsterone-only), and the pilocarpine-only injection group (pilocarpine-only).
Mice were anesthetized and transcardially perfused with heparinized saline, followed by 3.7% formaldehyde in phosphate-buffered saline (PBS). Brains were isolated and postfixed in the same fixative overnight at 4°C. Fixed frozen brains were sectioned coronally at 20 μm using a cryostat. Sections collected from single animals were used for histological analyses, such as cresyl violet staining and immunohistochemistry (n =6 mice in each experimental group).
For the DAB substrate reaction, sections were incubated with 3% H2O2 in PBS with 0.3% Triton X-100. CB was stained using the monoclonal anti-calbindin D28K antibody (1:3000, Sigma). The primary antibody was reacted overnight at 4°C. The secondary antibody, biotinylated anti-mouse IgG (Vector Laboratories, Burlingame, CA, USA), was reacted at room temperature for 1 h. During the procedure, all sections were washed with PBS between each step. We followed the manufacturer’s protocols for the M.O.M kit (Vector Labs). Visualization was performed using the Vectastain ABC-DAB system for 1 h at room temperature (Vector Labs).16
Sections were stained in 0.2% cresyl fast violet acetate for 5 min and rinsed in water. They were then rinsed in 95% alcohol and differentiated in cresyl violet differentiator [95% alcohol (90 mL), chloroform (10 mL), acetic acid (three drops)]. Sections were rinsed in absolute alcohol, cleared, and mounted on slides.16
Data are expressed as the mean ± standard error of the mean. Statistical comparisons were made using one-way analysis of variance (ANOVA), followed by Bonferroni’s post hoc tests (SAS, Cary, NC, USA). The level of significance was set at p<0.05 and 0.001.
Results
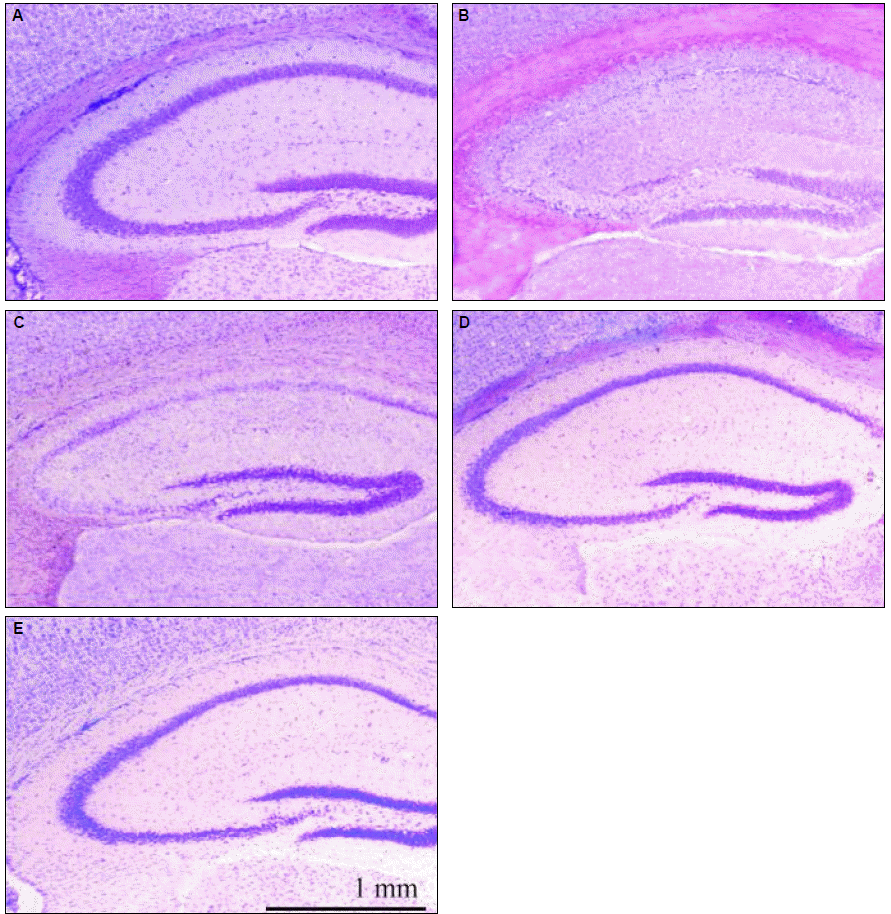
In the cresyl violet staining, hippocampal CA1 and CA3 pyramidal neurons were markedly decreased and morphologically-damaged neurons were observed in the pyramidal layer of the pilocarpine-only group. Hippocampal structures were relatively preserved in all other groups (Fig. 1). In the pilocarpine plus androsterone (200 mg) group, the structures of the hippocampal neurons in the CA1 and CA3 regions were as well preserved as the normal group (Fig. 1D).
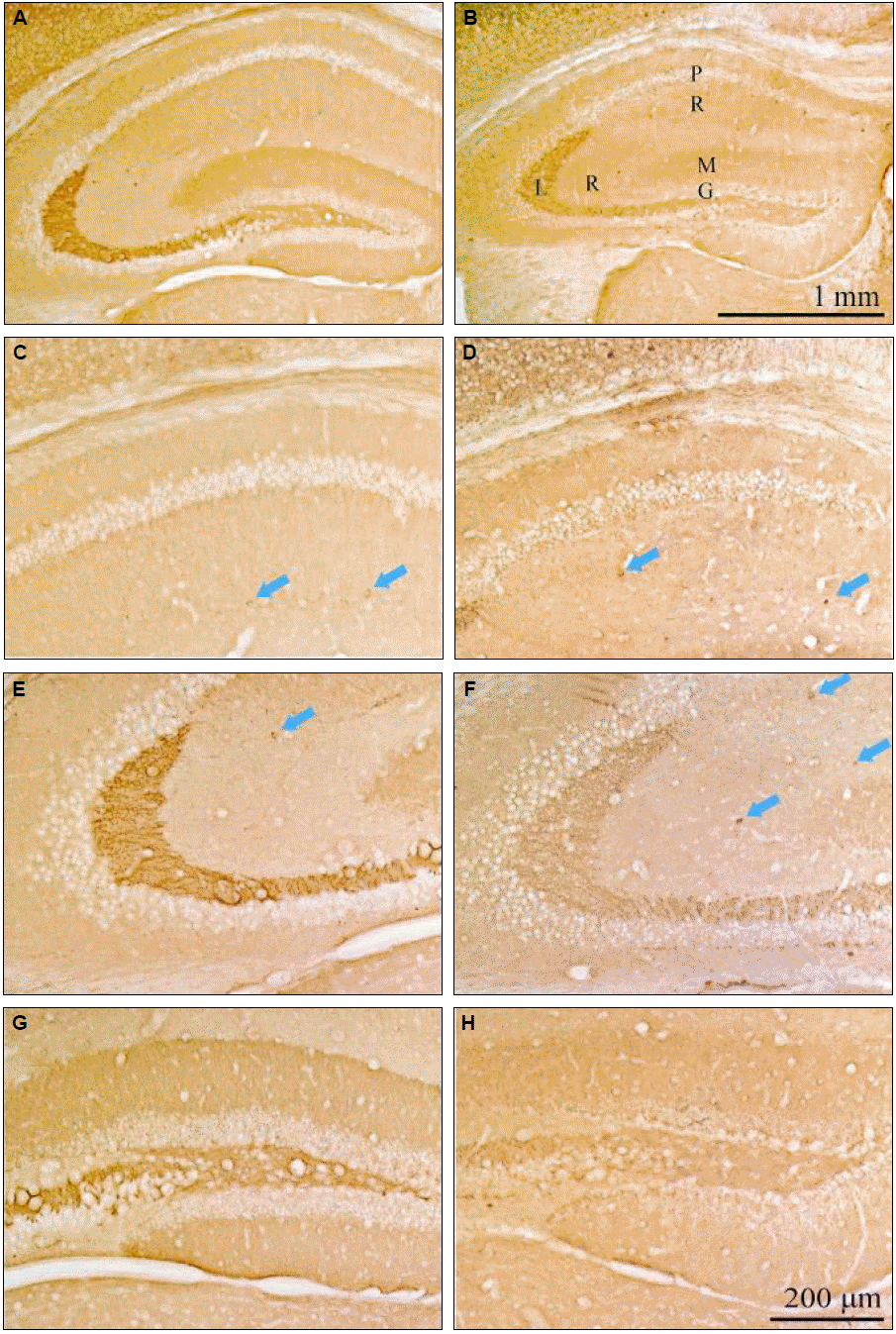
We evaluated the direct effect of androsterone on CB without SE and compared this with the normal and androsterone-only group (Fig. 2). CB-positive neurons were highly expressed in the stratum radiatum of the CA1 and CA3 region, stratum lucidum of the CA3 region, stratum molecular of the dentate gyrus (DG) and interneurons in the androsterone-only group (Fig. 2G and H).
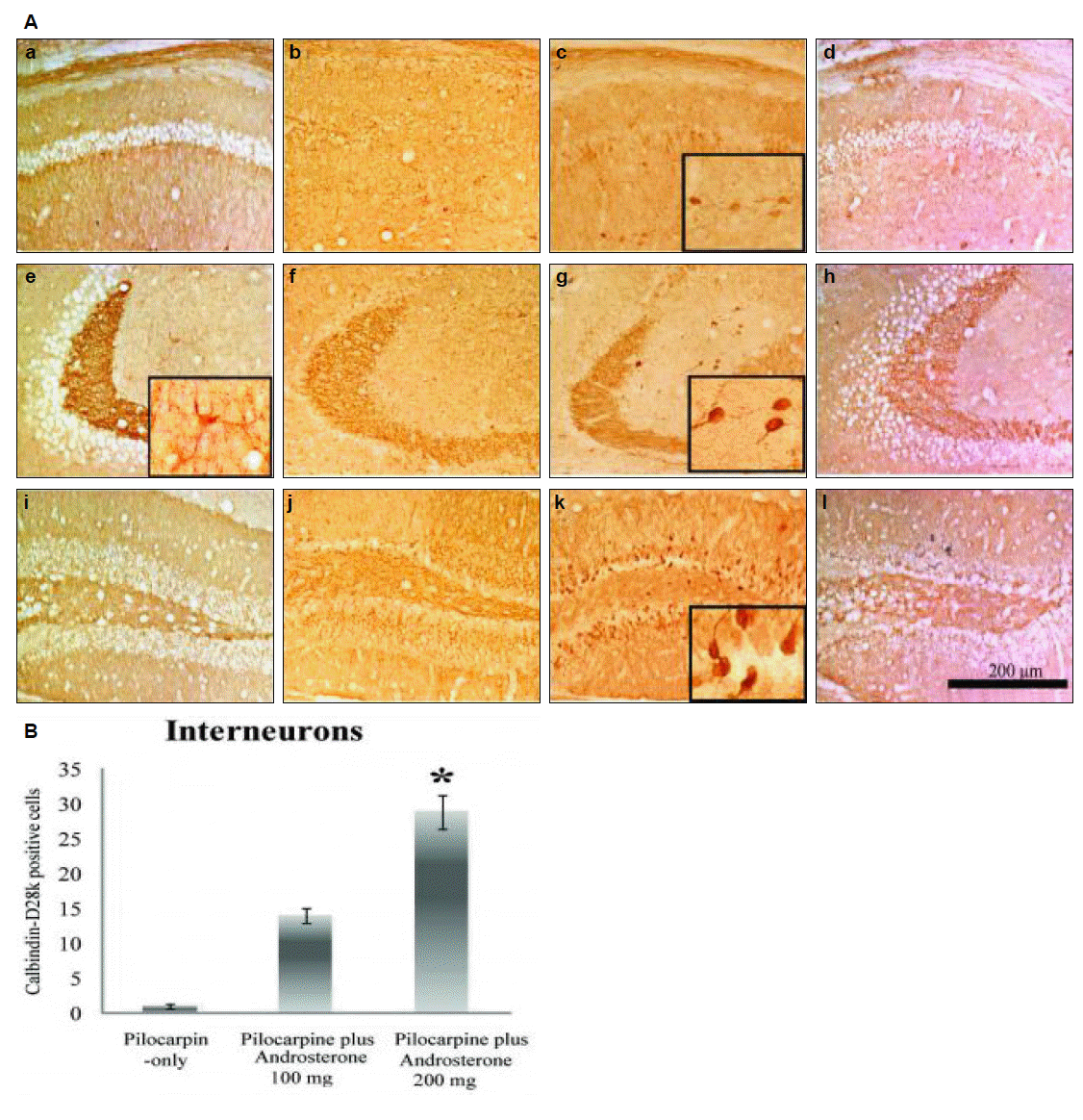
We also compared CB changes after pilocarpine-induced SE. CB-positive cells were increased in all subfields of hippocampus in the pilocarpine plus androsterone group compared to the pilocarpine-only group (Fig. 3). In particular, CB-positive cells increased dose-dependently in the pilocarpine plus androsterone group, the stratum radiatum of the CA1 and CA3 region, the stratum lucidum of the CA3 region (Fig. 3D to I), and the stratum molecular of the DG (Fig. 3J to L). In the pilocarpine-only group, CB-positive cells were dramatically reduced in the entire hippocampus. Especially in the stratum radiatum of the CA1 and CA3 regions, expression of CB-positive cells, such as interneurons, was reduced in pilocarpine-only group compared to the normal control group. Additionally, the CB staining intensity was decreased in the stratum lucidum of the CA3 region (Fig. 4).
As shown in Fig. 4, quantification of CB-positive interneurons in the stratum radiatum of the CA1 and CA3 regions was performed in the pilocarpine-only and pilocarpine plus androsterone groups. CB-positive cells in the pilocarpine-only group were not present; however, in the pilocarpine plus androsterone group, the number of CB-positive interneurons increased significantly compared to the pilocarpine-only group
The number of CB positive cells was 1±0.4 cells/mm3 in pilocarpine-only group, 14±1.1 cells/mm3 in pilocarpine plus androsterone 100 mg group and 29±2.5 cells/mm3 in pilocarpine plus androsterone 200 mg group (p<0.001) (Fig. 4B). In addition, the number of CB-positive interneurons increased dose-dependently. Finally, in the stratum lucidum of the CA3 region, the CB staining intensity in the pilocarpine plus androsterone group increased.
Discussion
Our results show a decrease in CB-positive cells in the stratum radiatum of the CA1 and CA3 regions of the hippocampus after pilocarpine induced-SE in the pilocarpine-only group. In contrast, androsterone treatment after pilocarpine-induced SE induced CB expression more than the pilocarpine-only group. CB-positive cells were detected more often in the stratum radiatum of the CA1 and CA3 regions in the androsterone plus pilocarpine group, and the morphology of neuronal cells was well preserved compared to the pilocarpine-only group. In the androsterone-only group, there tended to be more CB-positive compared to the normal control group.
Calcium influx and intracellular Ca2+ play role in several important functions, including production of action potentials, neurotransmitter release, cell-to-cell interactions, and neuronal plasticity in the central nervous systems. It has been reported that the failure to regulate intracellular Ca2+ concentration induces excessive Ca2+ influx into neuronal cells, resulting in a neuronal cell death cascade.17 In epilepsy, abnormal Ca2+ levels are found both in vitro and in vivo.18–20 Ca2+ levels were significantly and chronically elevated during bicuculline-induced epilepsy in vitro. Bicuculline, the GABAA receptor antagonist, also induced neuronal cell death.21,22 Therefore, Ca2+ is considered to play an important role in epileptogenesis.
CB is an intracellular calcium-binding protein, and functions in Ca2+ buffering systems. Therefore, CB may protect neuronal cells against in excessive Ca2+, including subpopulations of interneurons and the GABAergic character of a comparatively large group of non-pyramidal cells in the hippocampus.7,23,24 Hence, CB may regulate excitation of neuronal cells between excitatory and inhibitory neuronal cells. Previous studies found that decreased CB expression was observed in epileptic conditions.13 In addition, it was reported that some steroids, such as testosterone and estrogen, stimulated calbindin synthesis14,15 to regulate development of sexually dimorphic structures.18,19 Calbindin can modulate depolarization of dopaminergic cells,25 which is related to hippocampal cell death.26
Our finding of decreased numbers of CB-positive cells after pilocarpine treatment suggest that decreased CB is related to calcium homeostasis, and could be a major susceptibility to excite toxicity. The increase in CB after pilocarpine plus androsterone treatment suggests that androsterone modulates expression of calcium-binding proteins in neuronal cells during SE, which occurs in response to abnormal conditions such as excessive Ca2+. Additionally, the pyramidal cells in the hippocampus were preserved well in the pilocarpine plus androsterone group compared to the pilocarpine-only group. Taken together, our data suggest the increase in CB expression mediated by androsterone is related to neuronal cell protection in SE.
In conclusion, our observations suggest that the androsterone-mediated increase in CB-positive cells plays a role in buffering excess Ca2+ and protects neuronal cells. Androsterone may modulate Ca2+ homeostatic mechanisms in epilepsy and function in neuroprotection against excess Ca2+.